
The paper A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber has been published in the journal Current Biology by Lida Xing, Ryan C. McKellar, Xing Xu, Gang Li, Ming Bai, W. Scott Persons IV, Tetsuto Miyashita, Michael J. Benton, Jianping Zhang, Alexander P. Wolfe, Qiru Yi, Kuowei Tseng, Hao Ran, Philip J. Currie.1

Something which many will consider not worth noting for a few reasons but which is important to note as you read science papers is that Xing et al., do not merely write in a scientific manner of reporting just the facts ma’am. Thy do not merely say “We found thus and such” but they seek to build a narrative by seeking to (force?) fit their observations within the parameters of the theory of evolution. That is to say that they interpret evidence according to their evolution worldview-philosophy.
The paper states the following within that context:
…evolution of coelurosaurian integumentary structures…models of feather evolution…evolutionary developmental pathway of feathers…Within the evolutionary developmental model of feathers 2…intermediate between stages…the evolutionary pathway for feathers…Cytological observations of barbule development along the barb vane ridge support the evolutionary coupling of barbs and barbules3…the lineage leading to birds…evolutionary patterns…the context of bird origins…the origin of birds…enriched models of morphological character evolution that help explain major evolutionary transitions in key clades such as theropods, including birds…feather evolution…
There are also some weasel words such as “may have been through…may represent…evolutionary patterns remain difficult to interpret…taxonomic placement is uncertain…suggests either…or…”

Imaginative illustrations: an evolutionist’s best friend.
This all pertains to what is said to be a coelurosaur dinosaur which has been designated “DIP-V-15103.” Note that coelurosaurs is a very broad group of theropods which includes T. rex and birds.
Note that, the identification of the dinosaur is next to simply bypassed in the article. What was found is a disembodied tail and the article more so than not refers to “the feather bearer” in fact, note the big “IF” within the sentence, “If DIP-V-15103 indeed represents a juvenile coelurosaur tail” meaning that they are only speculating that it does (and making the thousands of articles which report something which the paper does not fake news). The sentence even states that “the feathers most likely characterize adult plumage; however, there is some room for uncertainty.” Ryan McKellar confirmed to me that the “If” regards the status as “juvenile” yet, when I replied thusly he never wrote back, “Thank you so much. That really helps because it seemed that the paper does not do much detailed arguing (or, evidencing) for as to why it is thought to be a coelurosaur.”
Of course, the paper states that which it wills at any given time such as that within amber was found the “feathered tail of a non-avialan theropod preserved in mid-Cretaceous (rv99 Ma) amber from Kachin State, Myanmar.”4 Another statement is “pristine feathers in direct association with a putative juvenile coelurosaur” (emphasis added for emphasis). And “the distribution and shape of the feathers only strongly supports placement crownward of basal coelurosaurs, such as tyrannosaurids and compsognathids.”
So, it is “adult plumage” or “belonged to a juvenile” and perhaps was or was not a coelurosaur. Frankly, they seem to all but simply assume that it was a dinosaur due to the evolutionary dogma de jour which disallows them from concluding that it was a bird.
Makes me wonder about how such papers may be subjected to the document hypothesis whereby one of the authors, this one had 14, asserts certainty while others admit uncertainty.
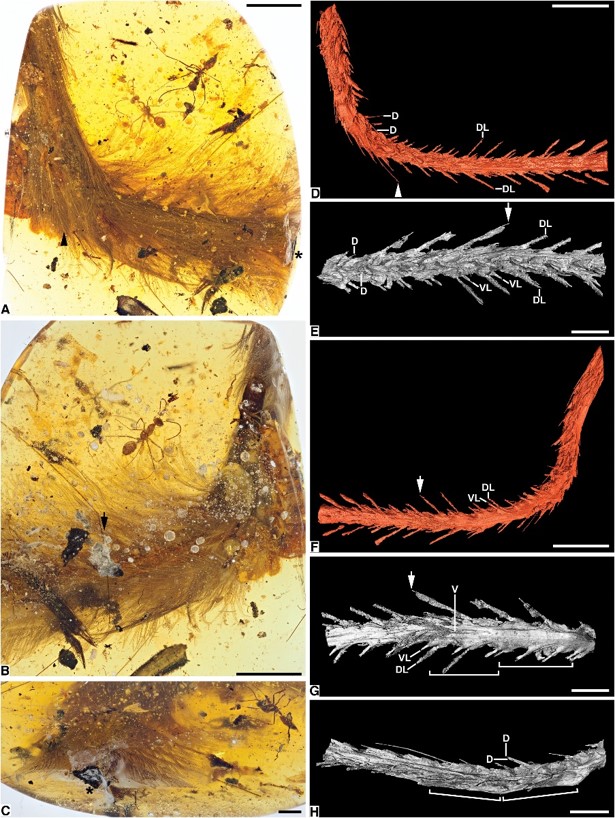
The paper notes, “The tail within DIP-V-15103 is visible to the naked eye as an elongate and gently curved structure (length = 36.73 mm). A dense covering of feathers protrudes from the tail…tail section contains at least eight full vertebrae and part of a ninth…was likely a relatively long tail.”

This is said to come “two decades since the discovery of feathered dinosaurs”5 which is to claim much more than the evidence represents: for elucidations of such claims see the following:
Is the feathered dinosaur “Anzu wyliei” the “chicken from hell”?
Atheist and Darwinian Science and Story Telling, part 8 of 9
Atheism and Archaeopteryx
Various articles on Alan Feduccia
Note that the paper notes that the amber and thus the specimen are circa 99 million years old and yet, “The open, flexible structure of these feathers is more analogous to modern ornamental feathers than to flight feathers” (emphasis added for emphasis) which goes to show that they have changed little, if at all, in the alleged 99 Ma.

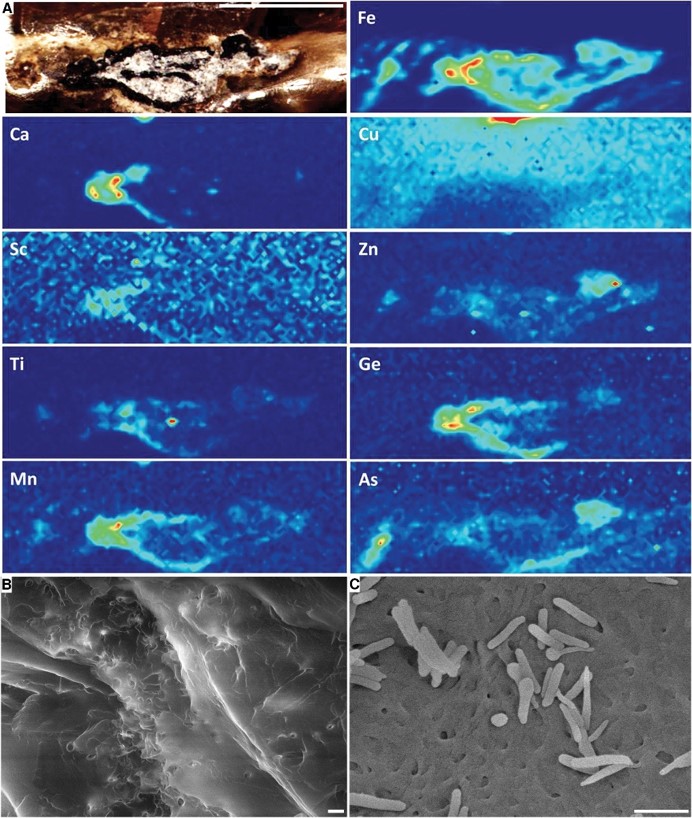
It is also noted that this 99 Ma amber contains soft tissue:
Beneath the feathers, carbonized soft tissues…suggests that vestiges of primary hemoglobin and ferritin remain trapped within the tail…Soft tissues—presumably muscles, ligaments, and skin—are visible…These tissues have largely been reduced to a carbon film, retaining only traces of their original chemical composition…the mummified external appearance of the skin…soft tissues have a density insufficiently different from the partially replaced skeletal elements to permit X-ray imaging and virtual dissection of osteology alone. Consequently, many diagnostic and comparative osteological details remain obscured. However, two vertebrae are clearly delineated ventrally.
Lastly, I will simply related some of the specific speculations:
…the barbule pattern in DIP-V-15103 may represent a highly derived and potentially experimental character state unrelated to the avian lineage…
models of feather evolution.6 However, three-dimensional feather morphology and evolutionary patterns remain difficult to interpret…
its vertebral profiles and estimated length rule out avebrevicaudan birds, oviraptorosaurs, and scansoriopterygians—lineages generally characterized by a short caudal series with subequal centra7, with the exception of Epidendrosaurus…
The branched feathers have a weak pennaceous arrangement of barbs consistent with non-avialan coelurosaurs, particularly paravians…
Although the feathers are somewhat pennaceous [having a stalk or quill], none of the observed osteological features preclude a compsognathid8 affinity [a genus of small theropod dinosaurs]…
can be confidently excluded from Pygostylia [a group of avialans: a clade of flying dinosaurs]; in addition, it can likely be excluded from the long-tailed birds…
DIP-V-15103 does not show the splaying of large pennaceous rectrices observed alongside the posteriormost caudals of long-tailed birds 9…
Amber preservation suggests that the tail fans and fronds preserved in paravians are not merely a taphonomic artifact of compression…
the feather bearer may not have conformed to the molt patterns found in modern birds…
The paired feather arrangement is similar to rectrices in modern birds…
the feather bearer would likely have been incapable of flight…
The feathers of DIP-V-15103 display exquisitely preserved barbules…
the feathers are comparable to the contours of many modern birds…
Within the evolutionary developmental model of feathers10, DIP-V-15103 appears to be intermediate between stages IIIa (rachis with naked barbs) and IIIb (barbs with barbules, lacking a rachis), but it does not exactly fit stage IIIa+b (rachis with barbs bearing barbules)…
suggests either a greater diversity of tail plumage in coelurosaurians than previously suspected or a simplified form of more derived pennaceous feathers in DIP-V-15103…
The contiguous barbule branching along the rachis probably occurs along the barb vane ridge leading to the apicalmost barb…
DIP-V-15103 suggests that non-avialan theropods had a greater variety of feather forms than predicted from developmental phenotypes in modern feathers 11…
~~~~~~~~~~~~~~~~~~~~~~~~
A plea: I have to pay for server usage and have made all content on this website free and always will. I support my family on one income and do research, writing, videos, etc. as a hobby. If you can even spare $1.00 as a donation, please do so: it may not seem like much but if each person reading this would do so, even every now and then, it would add up and really, really help. Here is my donate/paypal page.
Due to robo-spaming, I had to close the comment sections. However, you can comment on my Facebook page and/or on my Google+ page. You can also use the “Share / Save” button below this post.
- 1. Xing et al., A Feathered Dinosaur Tail with Primitive Plumage Trapped in Mid-Cretaceous Amber, Current Biology (2016), http://dx.doi.org/10.1016/j.cub.2016.10.008
- 2. Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306.
- 3. Multiple references:
Alibardi, L. (2006). Cells of embryonic and regenerating germinal layers within barb ridges: implication for the development, evolution and diversification of feathers. J. Submicrosc. Cytol. Pathol. 38, 51–76.Alibardi, L., and Sawyer, R.H. (2006). Cell structure of developing downfeathers in the zebrafinch with emphasis on barb ridge morphogenesis. J. Anat. 208, 621–642.
- 4. Grimaldi, D.A., Engel, M.S., and Nascimbene, P.C. (2002). Fossiliferous Cretaceous amber from Myanmar (Burma): its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 3361, 1–72.
- 5. Multiple references:
Ji, Q., and Ji, S.-A. (1996). On the discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds. Chin. Geol. 233, 30–33.Qiang, J., Currie, P.J., Norell, M.A., and Ji, S. (1998). Two feathered dinosaurs from northeastern China. Nature 393, 753–761.
Chen, P.J., Dong, Z.M., and Zhen, S.N. (1998). An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391, 147–152.
- 6. Multiple references:
O’Connor, J.K., Chiappe, L.M., Chuong, C.M., Bottjer, D.J., and You, H. (2012). Homology and potential cellular and molecular mechanisms for the development of unique feather morphologies in early birds. Geosciences (Basel) 2, 157–177.Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306.
Xu, X., Zhou, Z., and Prum, R.O. (2001). Branched integumental structures in Sinornithosaurus and the origin of feathers. Nature 410, 200–204.
Prum, R.O., and Dyck, J. (2003). A hierarchical model of plumage: morphology, development, and evolution. J. Exp. Zoolog. B Mol. Dev. Evol. 298, 73–90.
Xu, X., Zheng, X., and You, H. (2010). Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature 464, 1338–1341.
Xu, X., Zhou, Z., Dudley, R., Mackem, S., Chuong, C.M., Erickson, G.M., and Varricchio, D.J. (2014). An integrative approach to understanding bird origins. Science 346, 1253293.
Chen, C.-F., Foley, J., Tang, P.C., Li, A., Jiang, T.X., Wu, P., Widelitz, R.B., and Chuong, C.M. (2015). Development, regeneration, and evolution of feathers. Annu. Rev. Anim. Biosci. 3, 169–195.
- 7. Multiple references:
Zhang, F., Zhou, Z., Xu, X., Wang, X., and Sullivan, C. (2008). A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. Nature 455, 1105–1108.O’Connor, J.K., and Zhou, Z. (2013). A redescription of Chaoyangia beishanensis (Aves) and a comprehensive phylogeny of Mesozoic birds. J. Syst. Palaeontology 11, 889–906.
Persons, W.S., Currie, P.J., and Norell, M.A. (2014). Oviraptorosaur tail forms and functions. Acta Palaeontol. Pol. 59, 553–567.
- 8. Currie, P.J., and Chen, P.J. (2001). Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Can. J. Earth Sci. 38, 1705–1727.
- 9. O’Connor, J.K., Sun, C., Xu, X., Wang, X., and Zhou, Z. (2012). A new species of Jeholornis with complete caudal integument. Hist. Biol. 24, 29–41.
- 10. Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306.
- 11. Multiple references:
O’Connor, J.K., Chiappe, L.M., Chuong, C.M., Bottjer, D.J., and You, H. (2012). Homology and potential cellular and molecular mechanisms for the development of unique feather morphologies in early birds. Geosciences (Basel) 2, 157–177.Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Exp. Zool. 285, 291–306
Chen, C.-F., Foley, J., Tang, P.C., Li, A., Jiang, T.X., Wu, P., Widelitz, R.B., and Chuong, C.M. (2015). Development, regeneration, and evolution of feathers. Annu. Rev. Anim. Biosci. 3, 169–195.