
Herein we continue, from part 1, considering Alan Feduccia who is the S. K. Heninger Distinguished Professor (Emeritus) within University of North Carolina at Chapel Hill’s Biology Department.
Specifically, his article “Bird Origins Anew,” The Auk – An International Journal of Ornithology, Vol. 130 No. 1 January 2013 AD (bold and underlining emphasis has been added for emphasis).
Alan Feduccia notes:
At the Society of Vertebrate Paleontology’s fall 1996 meeting at the American Museum of Natural History, a small black-and white photograph of a newly discovered specimen purported to be a feathered dinosaur was pinned on a bulletin board, just months following discovery.
Figure 2With no light or scanning electron microscopy or other biological assay, a pen-and-ink of the specimen appeared on the front page of the New York Times as important new support for the theory that birds are derived from dinosaurs.
Following the current paleontological thought, a filamentous line along the back of the small compsognathid dinosaur Sinosauropteryx (“Chinese reptile wing”) was described by a Chinese invertebrate paleontologist as representing protofeathers, with no evidence to support the supposition.

In fact:
…there was never any evidence that these filaments represented anything other than a skeletal meshwork of collagen fibers supporting a lizard-like frill, scalloped along the tail as in the modern Central American basilisk lizards (Feduccia et al. 2005, Lingham-Soliar et al. 2007).
As it turns out “the fibers are internal” and “not to mention the presence of scales on the parts of the body (Martin and Czerkas 2000, Lingham-Soliar 2013; Fig. 1).”

Figure 1 states:
Specimen of the “Chinese reptile wing” or “downy dino” Sinosauropteryx prima (length 68 cm), described, with no evidence, as having a pelt of protofeathers…In all specimens examined, there is a clearly marked outline of the body (here shown with dotted line in the specimen and closeup of neck region below), showing that the filamentous zone is not external, but below the skin level, most likely a zone of collagen supporting a mid-dorsal, lizardlike frill (Lingham-Soliar et al. 2007; photo courtesy of John Ruben)…Decomposing collagen fiber bundles in the hypodermis of a dolphin…Plume-like patterns of some of the fibers along the edges of the decomposing tissue.
Many groups of fibers have narrow points of attachment (bottom arrow) that broaden before tapering again (due to degradation), giving the flame-like shape…Thick fiber bundles showing disorganization of the finer fibers and bundles to produce overlapping featherlike patterns…Detail of large fiber bundles showing component fibers during degradation, producing branching patterns among the filaments.
As is the case with Alan Feduccia’s examination of Archaeopteryx (see here for some details on this issue):
…no convincing evidence is presented that the “fibers” on the matrix represent feathers, and the carbonaceous markings on the ulna thought to be some form of ulnar quill nodes appear almost randomly ordered and do not remotely resemble those of modern birds.
This is even though “a late Cretaceous ornithomimosaur reported not only with body feathers, but with wing feathers that were used only for display (Zelenitsky et al. 2012).” Indeed, the artist is the Darwinian evolutionist’s best friend as whenever evidence is lacking, one can always hire an artist to paint, sculpt, computer animate the Darwinian tall tale mythical story. Find one such example of Gigantoraptor here where it is admitted the that fossil did not have feathers but the reconstruction does.
He also writes:
Considering that advanced avian wings were present in the Jurassic, trying to understand the origin of avian wings and remiges by studying Late Cretaceous theropod dinosaurs seems to stretch credulity.
Too, if their interpretation is true, it would be the only known case of such a phenomenon, therefore demanding the highest level of evidence, which is simply not available…As Philip Gingerich has aptly noted, “The problem is that we expect too much of morphology in asking it to tell us the genealogy of organisms as well as what they look like” (Gingerich 1988:628)…
“Why should we accept as convincing a method of systematic analysis that for ideological reasons trivializes all data sets other than morphology? We can do better!” (Dodson 2000:504).
Moreover:
McKellar and colleagues recently reported a variety of feather material in Late Cretaceous Canadian amber as being of dinosaur origin (McKellar et al. 2011, 2012), but careful critique of their microscopic techniques, morphological comparisons, and potentially incomplete analysis of other biological materials by Smithsonian feather expert Carla Dove and colleague Lorian Straker suggested that there was nothing from the amber specimens that did not conform nicely to modern avian feathers or their component parts and that the authors did not convincingly rule out other biological materials or microscopic artifacts (Dove and Straker 2012).
Also, “a popular misconception is that ulnar quill nodes are directly correlated with flight ability.”
Feduccia points out that “Today, the field of origins is stuck in a rut of circularity because all conclusions are based on the fact that ‘birds are living dinosaurs’” the results of which are, for example, the following assumptions or rather, assertions:
Birds are dinosaurs, thus Archaeopteryx is an earthbound dinosaur; flight originated from the ground up; ground-dwelling birds that resemble dinosaurs, but with true avian feathers, are feathered dinosaurs; any filamentous structures in the dinosaurian fossil record are protofeathers; and the four-winged gliders with true avian wings must be four-winged feathered dinosaurs.
Faulty pop theories are being applied even to non-bird flying animals:
…although a revival of the ground–up flight model for pterosaurs has been completely falsified (Feduccia 2012b:245–247), the idea still persists today, and is now even applied to bats (Kaplan 2011), despite a mountain of evidence to the contrary (Norberg 1990).
Here is an interesting comment on artistic reconstructions. Now, I refer to these as transconstructions because a reconstruction is meant to do just that re-construct as in back to its original. However, what many scientists are doing is taking whole skeletons or even mere fragments and transforming them into a construction that is made in their own image that is, the image they wish to see as a final product to prove their pop theory de jour:
…most Chinese fourwinged gliders are reconstructed as cursorial fly-swatters (Fig. 2), evolving flight from the ground up.

Moreover:
Aside from the phylogenetic context, the life reconstructions of these four-winged gliders as terrestrial cursors (Fig. 2) is clearly erroneous. They all have elongate hindlimb flight feathers that would have been a hindrance in terrestrial locomotion, and they were incapable of more than occasional terrestrial journeys: clear evidence indicates that these early birds were arboreal climbers and gliders (Alexander et al. 2010, Burnham et al. 2011). Yet the theme of the theropod origin of birds is that all things avian come from the ground up; there is hardly any other explanation for this continued egregious error…
Conversely, “Climbing four-winged gliders”:
…are reconstructed as earthbound theropods learning to fly from the ground up, with extended arms as though to reinvigorate Ostrom’s long-failed “fly-swatter model” for the origin of avian wings as exaptations for flight.
Here are some more points about faulty reconstructions:
…the microraptor Sinornithosaurus, typically reconstructed as an earthbound cursor, had elongate hind limb flight feathers, which would have impeded ground locomotion, and exhibits a well developed posterolateral bony flange and a strongly bowed outer metacarpal, making its hand better suited for support of primary feathers than that of Archaeopteryx (Paul 2002)…
Microraptors have been reconstructed in two distinctive models, the four-winged gliding model with sprawled hindlimb wings, by which it was originally described in Nature (Xu et al. 2003), and a dinosaurian bipedal model, or biplane model, by which it is reconstructed with the hindlimbs held beneath the body, incapable of sprawling, in other words, like a tiny T. rex.
The problem, of course, is that there is absolutely no reason the hindlimbs could not have been sprawled, as is the case in flying squirrels (Glaucomys spp.), flying lemurs (Dermoptera), etc., and even falling cats. Too, the sprawled model performs superiorly in wind-tunnel experiments (Alexander et al. 2010), most specimens are preserved with a sprawled posture, and the wing claws are adapted for trunk climbing (Burnham et al. 2011).
In addition, it would be difficult to imagine how selection could produce elongate, asymmetric hindlimb flight remiges by the most current paleontological reconstructions, in which the hindlimbs are held in flight beneath the body in obligate bipedal fashion, with elongate hindlimb wing feathers trailing behind, simply slicing through the air (Balter 2012).
See figure 2 above.
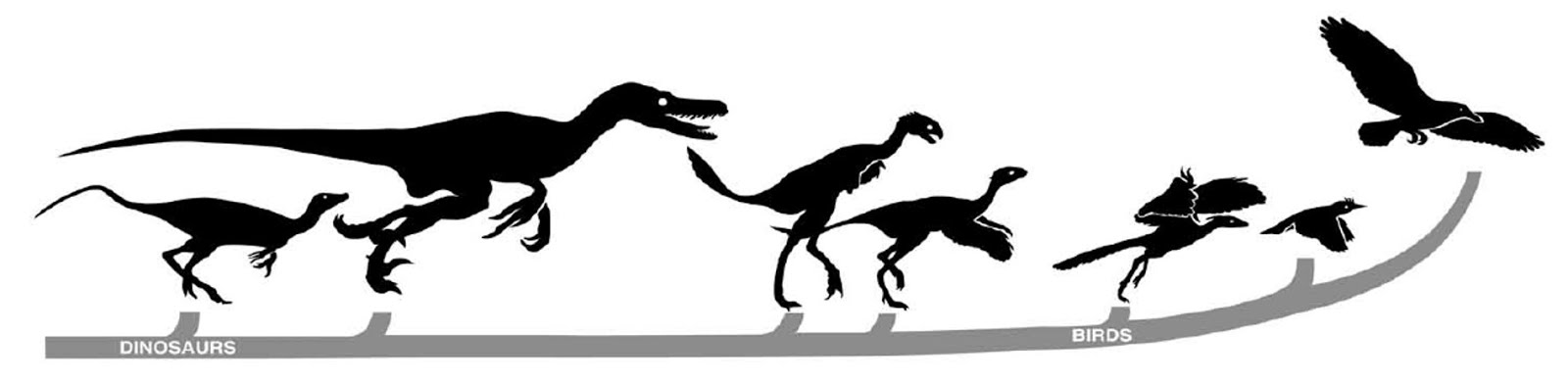
The notes to figure 6:
Topsy-turvy progression of fossil silhouettes associated with avian ancestry, displaying the current orthodoxy of bird origins, as interpreted by an article from National Geographic, but outrageously flawed. Harvard’s Alfred Romer would say: In discussing fossils, some notion of the geological time scale is necessary!
Censorship can also come in the form of simply ignoring data:
Most disturbing is that a massive, impeccably rendered cladistic analysis performed by the highly respected Polish paleontologist Maryańska (Maryańska et al. 2002; also see Lü et al. 2002), in which oviraptorosaurids clearly slotted as avians, is largely ignored by paleontologists and not even cited in a recent book on bird origins (Chiappe 2007)…
Another landmark paper never cited in recent paleontological literature is Cladistics and the Origin of Birds, by Frances James and John Pourtless (James and Pourtless 2009), in which they present two new cladistic analyses as well as an evaluation of all current hypotheses of bird origins.
Their conclusion, that presently “uncertainties about the hypotheses that birds are maniraptoran theropods are not receiving enough attention (p. 1),” was apparently sufficient incentive for the paper to be totally ignored. Lack of citation has become a common but disturbing mechanism of censorship (Feduccia 2009), and in addition to the above, you will find no citation in Chiappe’s book of the sophisticated work on flight origins by Ulla Norberg supporting the arboreal origin of flight (Norberg 1990; and numerous papers by Walter Bock [1986 and elsewhere]), or the extensive rebuttals to the existence of dinosaurian protofeathers by T. Lingham-Soliar (Lingham-Soliar 1999, 2001, 2003a, b, 2008, 2010a, b, 2011, 2012, 2013; Lingham-Soliar et al. 2007; Lingham-Soliar and Glab 2010; Lingham-Soliar and Plodowski 2010; Lingham-Soliar and Wesley-Smith 2008).
The filamentous band running down the neck on the small compsognathid dinosaur Sinosauropteryx, and on other theropods, ornithischians, and allies, are protofeathers. Yet no biological assays were applied to these structures, not even proof that they are hollow, a sine qua non for feather status. In fact, the best explanation for many of these structures is that they represent a skeletogenous matrix of collagen fibers supporting a lizard-like frill as in Sinosauropteryx (Lingham-Soliar 2003a, b; Feduccia et al. 2005; Lingham-Soliar et al. 2007), or other integumentary derivatives in varied archosaurs.
As noted, the preservational halos surrounding these fossils may be extraordinarily complex and may contain myriad structures, including preservationally degenerate and even macerated contour feathers that may be misidentified as protofeathers. If they were protofeathers they would be maladaptive, becoming wet or mucky. Successful young downy birds mature and abandon the downy state as quickly as possible.
When wet, baby Ostriches must seek shelter under the mother’s wing or risk hypothermia and eventual death. Despite the array of problems surrounding the identification of putative protofeathers, these structures are now conflated with true avian feathers in microraptors and other four-winged birdlike forms, which have true avian feathers (see 5 below [you can find fig 5 here]), causing further confusion. If an Ostrich head were retrieved from the Chinese Early Cretaceous fossil beds, it would no doubt be interpreted as having protofeathers of varied types, representing stages of feather evolution.
Another aspect is to keep promulgating debunked theories that were once the pop de jour theories—such as the entire theory of evolution, by the way:
Luis Chiappe, Kevin Padian, and Kenneth Dial are notably insistent on promoting the improbable cursorial model, the latter two recently extending it to bats, based on application of the long-debunked “biogenetic law” of Ernst Haeckel (Kaplan 2011, Balter 2012).
Another thing with which to contend is pop news report about science and also the ever present need to interpret evidence:
Xu et al. (2011), in an analysis in which Archaeopteryx was removed from Aves and slotted with deinonycosaurs. Nature News immediately headlined “Archaeopteryx no longer first bird” (27 July 2011 | Nature | doi:10.1038/news.2011.443), but shortly thereafter revised analysis by Lee and Worthy (2012) and Senter et al. (2012) reinstated the iconic Urvogel to its rightful place.
Such uncertainty illustrates that most cladograms hang by a spider’s thread, and if one were to eliminate the co-correlated gait characters and emphasize the uncoded innumerable avian features of four-winged microraptors and closely allied Jurassic Anchiornis and Xiaotingia, as well as the scansoriopterids, a completely different cladogram would emerge showing avian affinity.
As Clark noted years ago, “similarity lies in the eyes of the beholder, and…the particular hypothesis being advocated strongly colors perceptions of morphological resemblance” (Clark 1992:533).
In the next segment, we will consider Feduccia’s view on cladistics.
~~~~~~~~~~~~~~~~~~
References:
Alexander, D. E., E. Gong, L. D. Martin, D. A. Burnham, and A. R. Falk. 2010. Model tests of gliding with different hindlimb configurations in the four-winged dromaeosaurid Microraptor gui. Proceedings of the National Academy of Sciences USA 107:2972–2976Balter, M. 2012. Flying dinos and baby birds offer new clues about how avians took wing. Science 338:591–592.
Burnham, D. A., A. Feduccia, L. D. Martin, and A. R. Falk. 2011. Tree-climbing—a fundamental avian adaptation. Journal of Systematic Palaeontology 9:103–107.
Chiappe, L. M. 2007. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Wiley, New York
Dove, C. J., and L. C. Straker. 2012. Comment on “A diverse assemblage of Late Cretaceous dinosaur and bird feathers from Canadian amber.” Science 335:796
Feduccia, A., T. Lingham-Soliar, and J. R. Hinchliffe. 2005. Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. Journal of Morphology 266:125–166.
Feduccia, A. 2009. A colourful Mesozoic menagerie. [Review of Feathered Dinosaurs: The Origin of Birds by John Long. Oxford:Oxford University Press, 2009.] Trends in Ecology & Evolution 24:415–416.
Feduccia, A. 2012b. Riddle of the Feathered Dragons: Hidden Birds of China. Yale University Press, New Haven, Connecticut
James, F. C., and J. A. Pourtless. 2009. Cladistics and the origin of birds: A review and two new analyses. Ornithological Monographs, no. 66.
Kaplan, M. 2011. New theory on bat flight has experts a-flutter. Nature News Blog, 5 November. [Online.] Available at http://blogs.nature.com/news/2011/11/new_theory_on_bat_flight_has_e_1.html.
Lingham-Soliar, T. 1999. Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proceedings of the Royal Society of London, Series B 266:2367–2373.
Lingham-Soliar, T. 2001.The ichthyosaur integument: Skin fibers, a means for a strong, flexible and smooth skin. Lethaia 34:287–302.
Lingham-Soliar, T. 2003a. Evolution of birds: Ichthyosaur integumental fibers conform to dromaeosaur protofeathers. Naturwissenschaften 90:428–432.
Lingham-Soliar, T. 2003b. The dinosaurian origin of feathers: Perspectives from dolphin (Cetacea) collagen fibers.
Naturwissenschaften 90:563–567.Lingham-Soliar, T., A. Feduccia, and X. Wang. 2007. A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres. Proceedings of the Royal Society of London, Series B 274:1823–1829.
Lingham-Soliar, T., and J. Wesley-Smith. 2008. First investigation of the collagen D-band ultrastructure in fossilized vertebrate integument. Proceedings of the Royal Society of London, Series B 275:2207–2212
Lingham-Soliar, T. 2008. A unique cross section through the skin of the dinosaur Psittacosaurus from
China showing a complex fibre architecture. Proceedings of the Royal Society of London, Series B 275:775–780.Lingham-Soliar, T., and J. Glab. 2010. Dehydration: A mechanism for the preservation of fine detail in fossilized soft tissue of ancient terrestrial animals. Palaeogeography, Palaeoclimatology, Palaeoecology 291:481–487
Lingham-Soliar, T., and G. Plodowski. 2010. The integument of Psittacosaurus from Liaoning Province, China: Taphonomy, epidermal patterns and color of a ceratopsian dinosaur. Naturwissenschaften 97:479–486
Lingham-Soliar, T. 2010a. Dinosaur protofeathers: Pushing back the origin of feathers into the Middle Triassic? Journal of Ornithology 151:193–20
Lingham-Soliar, T. 2011. The evolution of the feather: Sinosauropteryx, a colourful tail. Journal of Ornithology 152:567–577.
Lingham-Soliar, T. 2012. The evolution of the feather: Sinosauropteryx, life, death and preservation of an alleged feathered dinosaur. Journal of Ornithology 153:699–711.
Lingham-Soliar, T. 2013. The evolution of the feather: Scales on the tail of Sinosauropteryx and an interpretation of the dinosaur’s opisthotonic posture. Journal of Ornithology 154: in press
Lü, J., Z. Dong, Y. Azuma, R. Barsbold, and Y. Tomida. 2002. Oviraptorosaurs compared to birds. Pages 175–189 in Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution (Z. Zhou and F. Zhang, Eds.). Science Press, Beijing
Martin, L. D., and S. A. Czerkas. 2000. The fossil record of feather evolution in the Mesozoic. American Zoologist 40:687–694.
Maryańska, T., H. Osmólska, and M. Wolsan. 2002. Avialan status for Oviraptorosauria. Acta Palaeontologica Polonica 47:97–116
McKellar, R. C., B. D. E. Chatterton, A. P. Wolfe, and P. J. Currie. 2011. A diverse assemblage of Late Cretaceous dinosaur and bird feathers from Canadian amber. Science 333:1619–1622.
McKellar, R. C., B. D. E. Chatterton, A. P. Wolfe, and P. J. Currie. 2012. Response to comment on “A diverse assemblage of Late Cretaceous dinosaur and bird feathers from Canadian amber.” Science 335:796.
Paul, G. S. 2002. Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore, Maryland
Xu, X., Z. Zhou, X. Wang, X. Kuang, F. Zhang, and X. Du. 2003. Four-winged dinosaurs from China. Nature 421:335–340.
Xu, X., H. You, K. Du, and F. Han. 2011. An Archaeopteryx-like theropod from China and the origin of Avialae. Nature 475:465–470
Zelenitsky, D. K., F. Therrien, G. M. Erickson, C. L. DeBuhr, Y. Kobayashi, D. A. Eberth, and F. Hadfield. 2012. Feathered non-avian dinosaurs from North America provide insight into wing origins. Science 338:510–514
~~~~~~~~~~~~~~~~~~~~~~~~
A plea: I have to pay for server usage and have made all content on this website free and always will. I support my family on one income and do research, writing, videos, etc. as a hobby. If you can even spare $1.00 as a donation, please do so: it may not seem like much but if each person reading this would do so, even every now and then, it would add up and really, really help. Here is my donate/paypal page.
Due to robo-spaming, I had to close the comment sections. However, you can comment on my Facebook page and/or on my Google+ page. You can also use the “Share / Save” button below this post.